

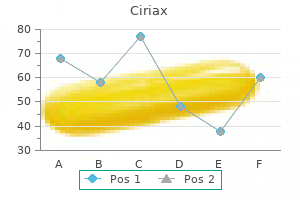
Ciriax dosages: 1000 mg, 750 mg, 500 mg, 250 mg
Ciriax packs: 30 pills, 60 pills, 90 pills, 120 pills, 180 pills, 270 pills, 360 pills
Purchase ciriax 750 mg mastercard
This kinase signals to not solely Akt but also a second kinase klebsiella antibiotic resistance mechanism ciriax 250 mg online buy cheap, mTor oral antibiotics for acne during pregnancy buy 250 mg ciriax visa, present in mTor complex 1 (mTorC1). All these responses could be anticipated to be useful for completion of viral infectious cycles. In fact, in every case that has been examined, virus infection has been noticed to activate signaling by way of Pi3k to Akt, and in lots of instances, mTorC1. The products of such genes can induce everlasting activation of cell proliferation, a process termed transformation, and sometimes acquisition of the flexibility to kind tumors in animals. Furthermore, the genomes of a big selection of viruses encode proteins that intervene downstream of Pi3k to preserve mTor exercise and consequently efficient translation (Chapter 11). Infection with a Particular Virus Modulates Multiple Signal Transduction Pathways Virus replica is invariably accompanied by alterations in more than a single signaling relay, usually with a quantity of pathways blocked and others stimulated. Concurrently, signaling cascades that govern other processes are modulated to support the reactions necessary for expression and replication of viral genomes and assembly of progeny virus particles. Infection by viruses with even comparatively easy genomes and mechanisms of reproduction that depend upon a minimal set of mobile systems and elements results in modification of a quantity of signaling pathways. Pi3k is activated following attachment to mobile receptors of adenoviruses, filoviruses, flaviviruses, influenza viruses, herpesvirus, and poxviruses, amongst others. Signaling initiated by the action of Pi3k ends in actin remodeling by way of activation of the small G proteins Rac and Cdc42. Inhibition of phosphorylation of Cas or production of a dominant-negative derivative of Rac inhibits adenovirus entry, emphasizing the significance of this mobile signaling pathway for environment friendly internalization. In this case, these processes rely upon signaling by way of Akt and focal adhesion kinase (Fak), and Pi3k is activated following clustering of lipid rafts and their related receptor protein tyrosine kinases (Rptks) in the plasma membrane. Such focus of receptors facilitates their activation by cross-phosphorylation, and likewise activates Mapk1 and -3. Such signal transduction not directly facilitates launch of the viral genomes into the cytoplasm by blocking diversion of endosomes containing virus particles to a nonproductive pathway. However, the ability of the mutant virus to form plaques on these cells was impaired (see the figure), as was induction of apoptosis when assessed by three totally different assays. In general, it seems that the more elaborate the strategy for viral reproduction, the higher the impression of an infection on signaling pathways. Furthermore, how radically cellular signaling systems are altered may even be decided by the origin and proliferation state of the host cell. Many human cells in routine use in the laboratory are derived from tumors (Chapter 2), and consequently are abnormal in many respects, together with unrestrained proliferation and everlasting activation of signaling circuits that promote cell growth and progression through the cell cycle. In contrast, in pure infections, many host cells proliferate solely slowly or are quiescent (withdrawn from the cell cycle). Successful virus reproduction in such cells is due to this fact likely to depend to a higher degree on activation of signaling pathways that management these processes than does copy in tumor-derived cell lines. The outcomes of recent purposes of those methods to comparison of uninfected and virus-infected cells recommend that the impact of explicit viruses on host cell signaling is even broader than previously appreciated. For example, comparison of the concentrations of phosphopeptides in uninfected, quiescent mouse fibroblasts and 18 h after infection with murine herpesvirus sixty eight (a gammaherpesvirus) recognized changes in 86% of the practically 2,500 unique peptides examined. This infection-induced distinction is much larger than that observed following exposure of cells to development factors (13%) or assaults, corresponding to injury to the genome or exposure of human cells to Salmonella (24%). Large-scale analyses of phosphoproteins in infected cells can even establish cellular substrates of signaling pathways which are important for virus replica. Subsequent functional evaluation of manufacturing of proteins with increased phosphorylation in infected cells established the necessary contribution of host proteins not previously implicated in reproduction of human immunodeficiency virus kind 1, notably a particular set of splicing proteins. Gene Expression Altered host cell gene expression is a common consequence of virus an infection. The influence of virus an infection on mobile gene expression and the mechanism(s) by which this process is altered differ with the strategies by which viral genes are expressed. Many of these proteins are described in previous chapters (Chapters 8, 10, and 11). Colors from yellow to blue and to red indicate decreases and increases, respectively. Moss, National Institute of Allergy and Infectious Diseases, National Institutes of Health. The very quick infectious cycle of poliovirus (and different picornaviruses; some eight h) may necessitate notably effective measures to forestall synthesis of mobile proteins. Indeed, when the concentrations of cellular proteins have been assessed early and late after vaccinia virus infection of human cells, 10% had been noticed to change considerably. Virus replica is dependent upon many secure mobile proteins, including ribosomal proteins and structural proteins of the cytoskeleton. However, it could even be necessary to maintain the production of a lot much less steady host proteins for optimal replica of specific viruses, even within the face of widespread inhibition of mobile gene expression. The impact of infection on patterns of mobile gene expression varies with virus. When combined with various types of bioinformatics evaluation, similar to classification of differentially expressed genes by their useful annotations (gene ontology analysis), the results of these descriptive research may help determine cellular gene merchandise and pathways that promote or counter virus copy, or that correlate with virus pathogenicity or the responses of individual hosts to infection (Box 14. Infection of quiescent, normal human fibroblasts by human adenoviruses is adopted by will increase or decreases of no less than 2-fold in expression of 10% of mobile genes. Many of those adjustments are associated with reentry of quiescent cells into the cell cycle, or help genome replication and expression. Transcription of a subset of genes repressed in contaminated cells, particularly interferon-sensitive and different genes associated with antiviral defenses, is inhibited by the viral E1B 55-kDa protein. Global reprogramming of the cellular translational panorama facilitates cytomegalovirus replication. The absorbance at 254 nm (shown) was monitored during assortment of gradient fractions. When mixed with genome-wide evaluation of mutations and adjustments in copy quantity, this approach has the potential to inform rationally based mostly personalised remedy. In a really uncommon subset of people contaminated with human immunodeficiency virus sort 1, reproduction of the virus (that is, synthesis of genomes) is undetectable by all but extraordinarily delicate assays. The number of elite controllers with gene expression profiles like these of uninfected persons was small (4 of the 12 elite controllers examined), but further research of the properties of their T cells could present clues in the search for a treatment for human immunodeficiency virus kind 1. In one complementary method to such conventional methods, the gene expression profiles of lung tissue of mice contaminated with influenza virus strains that differ in pathogenicity were in contrast. Note the clear separation of the profiles from uninfected persons and infected patients, and the dispersal of those from elite controllers in each clusters. Differences between the contaminated and mock-infected samples identified by microarray hybridization are proven for cytokine response, lipid metabolism, and coagulation genes. It has been advised that such a gene expression signature of extremely pathogenic influenza viruses may assist establish new therapeutics that target particular host proteins or pathways. Metabolism Host cells provide not solely the molecular equipment needed for synthesis of viral nucleic acids and proteins (at a minimum the translational machinery), but also the important constructing blocks, nucleotides and amino acids. Assembly of enveloped viruses also requires cellular membranes and the lipids from which these structures are constructed. Consequently, virus infection can lead to alterations within the pathways by which cells generate vitality from molecular fuels (catabolism), as well as those who make the precursors of nucleic acids, proteins, and membranes (anabolism).
Ciriax 250 mg
The bigger tubular airways embrace an epithelial layer lining the airway lumen; a submucosal layer containing connective tissue treatment for dogs eating chocolate ciriax 250 mg purchase free shipping, blood vessels antibiotics for sinus infection treatment 250 mg ciriax cheap with mastercard, lymphatics, and seromucinous salivary-type glands; a smooth muscle muscularis layer; and discontinuous cartilaginous rings offering structural integrity to the airways. The bronchial epithelium consists of pseudostratified ciliated columnar epithelial cells (which comprise nearly all of surface epithelial cells), mucin-secreting goblet cells, basal cells, and scattered neuroendocrine (Kulchitsky) cells. Neuroendocrine cells are positioned on the basement membrane, scattered among ciliated cells, and current in higher numbers in smaller bronchi. The submucosal layer lies beneath the epithelial layer and is fashioned by mesenchymal stroma containing variable amounts of fibrocollagenous tissue, elastic fibers, and seromucinous glands. These seromucinous salivary-type glands have a serous part with eosinophilic granular cytoplasm containing lysozyme and glycoproteins, as nicely as a mucinous component with more clear vacuolated cytoplasm containing mucin. The mucus secreted supplies safety to the lower respiratory tract as a half of the mucociliary escalator system. Bronchus-associated lymphoid tissues are lymphoid aggregates that are current throughout the submucosal tissue. In the bigger airways, the graceful muscle is organized in two sets of fibers in an opposing spiral orientation. The clean muscle bundles turn into less contiguous because the airways progress to the peripheral bronchioles. The C-shaped discontinuous cartilaginous rings of the mainstem bronchi are connected posteriorly by a clean muscle and elastic fiber�rich posterior membrane. Progressing more peripherally, the intrapulmonary bronchi have segmented into irregularly circumferential cartilage plates. As the bronchi department additional, the cartilage plates diminish and eventually remain only at the bifurcation point. Histologic Features the bronchus consists of a central lumen lined by respiratory 42 epithelium principally comprising pseudostratified ciliated columnar cells, goblet cells containing mucin, basal cells, and neuroendocrine (Kulchitsky) cells. The submucosal layer consists of fibrocollagenous loose connective tissue and elastic fibers with seromucinous glands that secrete mucus into the bronchial lumen. VanderLaan Bronchioles are the ultimate conductors of air that branch off from the bronchi. Bronchioles by definition are airways lower than 1 mm in diameter that lack cartilage and submucosal glands within their partitions. Bronchioles branch into terminal (membranous) bronchioles that are lined completely by ciliated columnar cells and nonciliated cuboidal Club cells. Terminal bronchioles further branch into respiratory bronchioles, that are lined by simple columnar to cuboidal epithelia that conduct air and participate in gasoline trade by way of alveoli of their walls. The lungs contain roughly 30,000 terminal bronchioles, every of which directs air to roughly 10,000 alveoli. The practical unit of the lung distal to the terminal bronchiole is known as the acinus, which consists of the respiratory bronchiole, alveolar ducts, and grape-like clusters of alveoli. A cluster of 3 to 10 terminal bronchioles, each with its corresponding acinus, is referred to as a lobule, certain together by an interlobular septum. The respiratory bronchiole usually offers rise to two or three generations of respiratory bronchioles, with rising numbers of alveoli budding from their partitions. These bronchioles then type alveolar ducts, tubular spaces sure entirely by alveoli, which terminate in alveolar sacs, the blind ends of the respiratory unit. As the airways of the acinus progressively branch out and diminish in caliber, the cuboidal epithelium transitions to more flattened 47 epithelial cells. Histologic Features Terminal (membranous) bronchioles branch off bronchi and are lined by ciliated columnar cells and nonciliated cuboidal Club cells. Respiratory bronchioles department off terminal bronchioles and are lined by simple columnar to cuboidal epithelia and alveoli. Respiratory bronchioles form alveolar ducts that are lined totally by alveoli and terminate in alveolar sacs. The acinus is the functional unit distal to the terminal bronchiole and consists of the respiratory bronchiole, alveolar ducts, and related alveoli. VanderLaan Sinchita Roy-Chowdhuri the lungs have a twin blood supply, comprising the pulmonary circulation (carrying deoxygenated blood from the right ventricle) and the bronchial circulation (carrying oxygenated blood from the left ventricle). The pulmonary circulation delivers blood from the best facet of the heart by way of progressively smaller branches of pulmonary arteries and arterioles to the small capillaries of the alveolar partitions where gas change occurs. The oxygenated blood is returned to the left atrium through pulmonary venules and veins. The bronchial circulation is a half of the high-pressure systemic circulation and supplies oxygen and vitamins to the bronchi and lung parenchyma. Pulmonary arteries and arterioles run alongside airways (bronchi and bronchioles) and, under normal circumstances, should have equal cross-sectional luminal diameters at any given level. The pulmonary veins usually have thinner muscular walls in contrast with pulmonary arteries, and sometimes their draining relationship to the interlobular septa and pleura could be identified with serial tissue sections. Elastic stains can be useful in highlighting the concentric elastic lamina of pulmonary arteries (the larger of which have each inner and external elastic laminae). Smaller arterioles, bronchial arteries, and pulmonary veins usually have only one (internal) elastic lamina. The histologic differentiation of pulmonary arterioles from 51 pulmonary venules could be difficult, but serial tissue sections can help differentiate the two. The lymphatics are typically inconspicuous vascular channels with out muscular vessel partitions and are lined by flat endothelial cells. The lymphatic distribution can turn into conspicuous in cases of pulmonary edema, lymphangitic carcinomatosis, or when examining lung parenchyma involved by sarcoid. Histologic Features Larger muscular pulmonary arteries have a tunica media layer comprising circularly oriented easy muscle cells bounded by inner and exterior elastic laminae. Smaller pulmonary artery branches and pulmonary arterioles have a thinner tunica media in contrast with bronchial arteries due to differences in strain between the two circulations. Pulmonary veins have comparatively skinny muscular partitions and have a single (internal) elastic lamina; nonetheless, morphologic arterialization can happen in circumstances of continual pulmonary venous hypertension. Lymphatic vessels are vague, however the channels can be found operating along bronchovascular bundles, in the interlobular septa, and in the pleura. VanderLaan Sinchita Roy-Chowdhuri Alveoli are a part of the terminal respiratory unit that consists of alveolar ducts and accompanying sac-like alveoli emanating from the respiratory bronchioles. Although each technology of airway is shorter and narrower than its precursor, the summed-up crosssectional surface space past the segmental bronchi is significantly excessive because of quite a few tiny alveoli that allow for gasoline exchange. The pores of Kohn, then again, are small holes found in the alveolar partitions. The cuboidal epithelium of the respiratory bronchiole abruptly transitions to the alveolar epithelium, which extends all through the alveoli. The alveolar wall consists of fifty eight pneumocytes and endothelial cells of the alveolar capillaries where fuel exchange happens between the inhaled air and the blood in the capillaries. The basement membranes of the pneumocytes and endothelial cells are sometimes fused. The alveolar interstitium is the connective tissue framework, which consists of collagen and elastin fibers, fibroblasts, myofibroblasts, pericytes, histiocytes, dendritic cells, Langerhans cells, mast cells, nerves, and nerve terminals. The alveolar epithelium, interstitium, and capillary endothelium make the air�blood barrier.
Buy ciriax 1000 mg with amex
In addition virus papiloma humano 500 mg ciriax discount with visa, Hcf binds to Zhangfei oral antibiotics for acne during pregnancy ciriax 750 mg buy online, a cellular protein that is a strong repressor of transcription. Zhangfei is synthesized in sensory neurons (a pure website of latency) however not in most other cell types. They can even inhibit apoptosis and intervene with expression of interferon genes, capabilities that would promote the survival of latently infected neurons. How then can transcription of immediate-early genes occur to provoke entry into the lytic cycle throughout reactivation from latency It is now clear that neurons damaged in this method exhibit large-scale adjustments in gene expression and, inside 2 to three h, characteristic features of neuronal degeneration. More lately, the mechanism of reactivation in latently infected trigeminal ganglion neurons of herpes simplex virus type 1-infected mice has been examined by exploiting viruses carrying reporters for expression of particular viral genes. When latent infections had been established (40 days later), the mice have been topic to hyperthermic stress to induce reactivation. Trigeminal ganglia had been removed 22 h thereafter, and the concentrations of infectious virus particles have been measured by plaque assay on cells in tradition. The ratios on the histograms present the number of mice optimistic versus the quantity examined. Inhibition of the Cellular Transcriptional Machinery Inhibition of mobile transcription in virus-infected cells presents a number of advantages. The essential participation of cellular transcriptional methods in the infectious cycles of most viruses thought of in this chapter precludes inactivation of this equipment. However, posttranscriptional mechanisms permit selective expression of adenoviral and herpesviral genes (Chapter 10). Furthermore, transcription of many cellular genes is inhibited following an infection with herpes simplex virus kind 1. Consequently, its cleavage by 3Cpro in poliovirus-infected cells seems to be a very efficient method to forestall transcription of all mobile genes. Two gene merchandise of the rhabdovirus vesicular stomatitis virus have been implicated in inhibition of cellular transcription. Given the large repertoire of molecular and biochemical activities displayed by components of the cellular transcriptional equipment, it appears likely that different unusual actions of those cellular proteins might be discovered in virus-infected cells. In distinction, poxviruses similar to vaccinia virus are reproduced completely within the cytoplasm of their host cells. Distinguishing intermediate from late genes has been troublesome: both are transcribed only after viral genome replication and their promoters share sequence similarities. In fact, development of a genome-scale map of those transcription units has been achieved only just lately (Box 8. It is due to this fact an axiom of molecular virology that every mechanism by which viral transcription models are expressed by cellular elements, or by which their activity is regulated, will prove to have a normal cellular counterpart. By definition, early genes are these transcribed prior to viral genome replication. This drawback is compounded by the close spacing of open reading frames and intensive read-through transcription from one gene into neighboring downstream genes. The dependence of late gene expression on dedicated initiation proteins was used to solve this drawback and to permit intermediate and late genes to be distinguished. These experiments relied on tightly inducible expression from the viral genome of the G8R gene, which encodes the protein essential for late transcription, under the management of the E. Subsequent viral gene expression is determined by viral genome replication and the ordered synthesis of viral proteins that permit sequential recognition of intermediate and late promoters. The viral genome additionally encodes several proteins that regulate elongation throughout transcription of late genes. This protein (Vitf2) is positioned in the nucleus of uninfected cells however is current in each the cytoplasm and the nucleus of contaminated cells. As a significant number of vaccinia virus genes encode proteins necessary for transcription, such dependence on a mobile protein must confer some benefit. An engaging risk is that interaction of the viral transcriptional machinery with a mobile protein serves to combine the viral reproductive cycle with the expansion state of its host cell. The identification of Vitf2 as a heterodimer of proteins which are produced in biggest quantities in proliferating cells is according to this hypothesis. In distinction, transcription of the overwhelming majority of vaccinia virus early genes does terminate at discrete websites, 20 to 50 bp downstream of particular T-rich sequences within the template. It is difficult to exaggerate the contributions of viral techniques to the elucidation of mechanisms of transcription and its regulation in eukaryotic cells. Perhaps even more importantly, efforts to elucidate the molecular foundation of regulatory circuits that are crucial to viral infectious cycles have established basic principles of transcriptional management. Furthermore, the mechanisms that allow sequential expression of viral genes are quite well established. The models for the person regulatory processes described in this chapter had been developed initially by using convenient and highly effective experimental systems. Nor can they handle such points as how transcription of specific genes can be coupled with replication of the viral genome. Many viral regulatory proteins perform a number of functions, a property that may confound genetic evaluation, and the examine of individual intracellular reactions, similar to binding of a protein to a specific promoter sequence, is technically demanding. Nevertheless, viral cis-acting sequences and regulatory proteins stay extra amenable to genetic analyses of their perform in the natural context than do their cellular counterparts. The metazoan Mediator co-activator complicated as an integrative hub for transcriptional regulation. Long distance relationships: enhancerpromoter communication and dynamic gene transcription. Role of an adenovirus E2 promoter binding consider E1A-mediated coordinate gene control. Identification of a practical promoter in the lengthy terminal repeat of Rous sarcoma virus. An inducible transcription factor that prompts expression of human immunodeficiency virus in T cells. Transactivation of a bovine papillomavirus transcriptional regulatory component by the E2 gene product. The role of Tat within the human immunodeficiency virus life cycle signifies a main effect on transcriptional elongation. Herpesviral latency-associated gene promotes assembly of heterochromatin on viral lytic-gene promoters in latent an infection. Adenovirus E1A prevents the retinoblastoma gene product from complexing with a cellular transcription factor. Characterization of the herpes simplex virion-associated issue responsible for the induction of genes. An adenovirus kind 5 early gene function regulates expression of different early viral genes. Transcription of a vaccinia virus late promoter template: requirement for the product of the A2L intermediate-stage gene. A cellular factor is required for transcription of vaccinia viral intermediate-stage genes.
Buy 1000 mg ciriax with amex
This method is a modern validation of the Hershey-Chase experiment described in Chapter 1 length of antibiotics for sinus infection order 1000 mg ciriax free shipping. Deletion mutations can be used to take away a complete gene to assess its position in replica antibiotic in food 1000 mg ciriax sale, to produce truncated gene merchandise, or to assess the features of particular segments of a coding sequence. Noncoding areas may be deleted to establish and characterize regulatory sequences corresponding to promoters. Substitution mutations, which may correspond to one or more nucleotides, are sometimes made in coding or noncoding regions. Included in the former class are nonsense mutations, in which a termination codon is launched, and missense mutations, during which a single nucleotide or a codon is modified, resulting in the synthesis of a protein with a single amino acid substitution. Substitutions are used to assess the roles of particular nucleotides in regulatory sequences or of amino acids in protein operate, corresponding to polymerase exercise or binding of a viral protein to a cell receptor. When mutations occur in open reading frames, they can be manifested as adjustments within the synthesized proteins. For example, a number of base adjustments in a particular codon may produce a single amino acid substitution, a truncated protein, or no protein. The terms "mutation" and "deletion" are sometimes used incorrectly, or ambiguously to describe alterations in proteins. In this textbook, these terms are used to describe genetic adjustments, and the phrases "amino acid substitution" and "truncation" are used to describe protein alterations. Genomes and Genetics 71 Introduction of mutagenized viral nucleic acid into cultured cells by transfection might produce one of a quantity of attainable outcomes. Reversion Analysis the phenotypes caused by mutation can revert in certainly one of two methods: by change of the mutation to the wild-type sequence or by acquisition of a mutation at a second site, either in the identical gene or a unique gene. Phenotypic reversion attributable to second-site mutation is named suppression, or pseudoreversion, to distinguish it from reversion on the unique website of mutation. Reversion has been studied for the reason that beginnings of classical genetic evaluation (Box three. In the trendy period of genetics, cloning and sequencing strategies can be used to show suppression and to determine the character of the suppressor mutation (see below). The identification of suppressor mutations is a robust software for studying protein-protein and protein-nucleic acid interactions. The allele specificity of second-site mutations provides proof for physical interactions amongst proteins and nucleic acids. Phenotypic revertants could be isolated either by propagating the mutant virus under restrictive situations or, within the case of mutants exhibiting phenotypes. Nucleotide sequence analysis is then used to decide if the original mutation continues to be current in the genome of the revertant. The presence of the unique mutation signifies that reversion has occurred by second-site mutation. The ultimate step is introduction of the suspected suppressor mutation into the genome of the original mutant virus to verify its impact. In genetic evaluation of viruses, mutations are made in vitro by a wide selection of strategies, all of which can introduce surprising adjustments. It is unlikely that an unlinked mutation with the identical phenotype would occur twice. If the mutation indeed causes the phenotype, the wild-type phenotype must be restored in the rescued virus. If the wild-type phenotype is restored (complemented), then the likelihood is high that the phenotype arises from the mutation. The merit of this methodology over marker rescue is that the latter exhibits only that unlinked mutations are most likely not the trigger of the phenotype. The second-site mutations that suppressed the replication defects had been localized to a selected area within the gene for large T antigen. These results indicated that a specific area of large T antigen interacts with the Ori sequence during viral genome replication. Nucleotide sequence evaluation of the genomes of two pseudorevertants demonstrated that they include base changes that restore the disrupted secondary structure. Diagrams of the area between nucleotides 468 and 534, which corresponds to stem-loop V (Chapter 11), are proven. These include, from left to proper, sequences of wild-type poliovirus kind 1, a mutant containing the nucleotide changes highlighted in orange, and two phenotypic revertants. For instance, one goal of gene therapy is to deliver a gene to sufferers who either lack the gene or carry faulty variations of it (Tables 3. The 7,000 monogenic human disorders, characterized by mutations in one gene, are especially amenable to viral gene therapy. Recombinant viruses carrying foreign genes can infect a higher percentage of cells and thus facilitate era of the specified population. A full understanding of the construction and performance of viral vectors requires knowledge of viral genome replication, a topic mentioned in subsequent chapters for chosen viruses and summarized within the Appendix. Genetically engineered viruses are also being used to deal with a broad variety of cancers, a field called viral oncotherapy (Box 3. Many tumor cells have defective innate immune signaling pathways and therefore are susceptible to viral lysis. Viruses used for this purpose may be made more selective for tumor cells in quite lots of methods. Another strategy to viral oncotherapy is to utilize nonhuman viruses, such because the poxvirus myxoma virus and the picornavirus Seneca Valley virus, which may infect cells in human tumors however not normal tissues. Design requirements for viral vectors embody the use of an applicable promoter; maintenance of genome measurement inside the packaging restrict of the particle; and elimination of viral virulence, the capacity of the virus to cause illness. Expression of foreign genes from viral vectors could also be managed by seventy four Chapter three Table 3. Such genes could be built directly into the viral genome or launched by recombination in cells, as described above (see "Introducing Mutations into the Viral Genome"). The viral vector genome usually carries deletions and typically extra mutations. Deletion of some viral sequences is commonly required to overcome the constraints on the scale of viral genomes that can be packaged in virus particles. Viruses from 9 completely different households (Adenoviridae, Picornaviridae, Herpesviridae, Paramyxoviridae, Parvoviridae, Reoviridae, Poxviridae, Retroviridae, and Rhabdoviridae) are at present in clinical trials to test their safety and anticancer properties. The genomes of many viruses have been modified to confer higher efficacy and specificity for tumor cells. A problem to the development of oncolytic viruses is the host antiviral immune response, which may blunt therapeutic efficacy. Several approaches have been used to address this downside, together with the substitution of structural proteins from totally different human or animal serotypes and the production of novel serotypes by chemical modification of virus particles. Different serotypes can be used when the patient is immune to the unique vector, due to either earlier infection or remedy. Viral structural proteins may be modified to bind proteins that are specific to the goal cells, conferring greater specificity for tumor lysis. For instance, many tumor genes are expressed at aberrantly excessive ranges; the promoters and enhancers liable for such excessive expression have been recognized and used to drive synthesis of viral genes encoding proteins that mediate in cell killing. Enhanced killing of tumor cells has additionally been achieved by inserting a gene in the viral vector that makes the cell more prone to destruction by medicine or immune therapies.
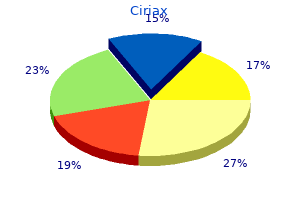
1000 mg ciriax generic with amex
Grossly antibiotic resistance arises due to quizlet ciriax 500 mg generic on-line, their appearance could differ bacteria energy source 750 mg ciriax generic otc, however undifferentiated sarcomas often manifest as a white-tan nodular mass with necrosis. Cytologic Features Mixture of malignant spindle cells, and occasional big cells, with areas of myxoid stroma and necrotic background. Histologic Features High-grade sarcoma with variable development patterns, brisk mitoses, occasional multinucleated tumor cells, necrosis, and hemorrhage. Morphologic subclassification includes undifferentiated spindlecell sarcoma, undifferentiated pleomorphic sarcoma, undifferentiated round cell sarcoma, undifferentiated epithelioid sarcoma, and undifferentiated sarcoma, not otherwise specified, designated by the predominant morphologic appearance of sarcoma cells. Grossly, these tumors are 327 usually tan-gray with distinguished necrosis and hemorrhage. Cytologic Features Primitive-appearing small to medium round blue cells with granular to nice chromatin and scant or inconspicuous cytoplasm. Histologic Features Most generally sheet-like association of small- to mediumsized spherical cells with fine chromatin, scant cytoplasm, and occasional neuroectodermal differentiation, including pseudorosettes. When neuroectodermal differentiation is prominent, Ewing sarcoma is usually termed primitive neuroectodermal tumor. Although histologically nonspecific, Ewing sarcoma is defined by recurrent genetic translocations (see below). Other primary websites, together with the lung, are attainable, though metastatic disease is more frequent and sometimes the presenting manifestation of the illness. Grossly, these tumors are poorly circumscribed, yellow-gray, and soft, with hemorrhage and necrosis. Histologic Features Organoid/nesting architecture, with delicate fibrovascular stroma separating similarly sized nests. Tumor cells are uniform and epithelioid, with distinguished cell borders, abundant, granular to clear cytoplasm, central uniform nuclei, with prominent nucleoli. Occasional rhomboid intracytoplasmic crystalline inclusions are apparent inside tumor cells. Ancillary Testing Electron microscopy sometimes shows membrane-bound lattice-like rhomboid crystals Periodic acid�Schiff highlights intracytoplasmic crystalline inclusions, that are sometimes focal. It much less commonly arises from head/neck, trunk, or visceral buildings, and when in the lung, it typically represents metastatic illness. Grossly, clear cell 335 sarcomas are lobulated, gray-white, with variable hemorrhage and necrosis, and uncommon pigmentation. Cytologic Features Highly cellular neoplasm with variably cohesive cells; eosinophilic, eccentric cytoplasm; spherical nuclei; and distinguished nucleoli. Pleomorphism, macronucleoli, and mitotic exercise are often much less outstanding than in metastatic melanoma. Neoplastic cells are comparatively uniform and spindled to epithelioid, with pale eosinophilic or amphophilic cytoplasm, vesicular nuclei, and outstanding nucleoli, though they are often as pleomorphic and mitotically energetic as metastatic melanoma. It rarely occurs outside the stomach cavity and, when current in the lungs, sometimes represents metastatic illness. Cytologic Features Primitive-appearing small to medium spherical blue cells with finely granular chromatin, indented nuclei, inconspicuous nucleoli, and scant cytoplasm. Histologic Features Sharply delineated, highly cellular nests inside a prominent collagenous or desmoplastic stroma are seen. The cells are usually uniform, small, and round, with hyperchromatic nuclei, scant cytoplasm, and inconspicuous borders. These tumors might arise in the extremities and are sometimes seen in visceral organs (most 345 generally liver) and, when in the lung, usually represent metastatic unfold. Cytologic Features Extrarenal rhabdoid tumors usually include dispersed, epithelioid cells with eccentric, spherical nuclei with prominent nucleoli, and abundant cytoplasm with perinuclear cytoplasmic density (rhabdoid morphology). Histologic Features Extrarenal rhabdoid tumors will typically grow in sheets and cords of discohesive "rhabdoid" cells with eccentric, vesicular nuclei, distinguished nucleoli, and perinuclear eosinophilic hyaline inclusions/globules. Smaller, round epithelial cells are additionally present with round nuclei and eosinophilic cytoplasm. Histologic Features Typically lobulated with abundant chondromyxoid stroma separated by fibrous stroma. Embedded within the stroma are ribbons and cords of enormous, physaliphorous (bubbly) cells with variable nuclear atypia as well as smaller, epithelioid cells with round nuclei. High-grade undifferentiated sarcoma juxtaposed to a conventional chordoma component defines dedifferentiated chordoma. Cytologic Features Round, uniform cells with moderate granular cytoplasm organized in small clusters or chords with ample myxoid background. Histologic Features Multinodular tumor with abundant gray-blue, hypovascular stroma separated by fibrous septae. Embedded throughout the stroma are small, barely elongated uniform cells with moderately plentiful cytoplasm, which combination in unfastened clusters, cords, or nests. They represent <1% of all primary lung tumors and 3% to 5% of extranodal lymphomas. Primary pulmonary lymphoma is defined as lymphoma confined to the lung with or with out hilar lymph node involvement on the time of diagnosis or up to 3 months thereafter. Secondary lung involvement by systemic B-cell lymphomas is by way more frequent than major disease. Histologically, all these disorders are similar to their counterparts in lymph nodes or other extranodal sites. Primary and secondary B-cell lymphomas of the lung are morphologically similar and may only be distinguished as 358 such by the clinical presentation. Moreover, these lymphomas could also be related to organizing pneumonia that may further complicate the histologic interpretation. When recent material is on the market, circulate cytometry might affirm the clonality of B cells by the detection of monotypic light chain expression or the expression of aberrant markers in T cells. Grossly, it consists of an ill-defined fleshy mass with preservation of the lung structure. A lesion could additionally be handled surgically whether it is peripheral or by statement or single-agent chemotherapy whether it is unresectable. Relapses might occur within the lungs, the stomach, salivary glands, and/or lymph nodes. Histologic Features Dense lymphoid infiltrate with a lymphangitic distribution along septa and bronchovascular bundles. Large coalescent lesions form a stable mass that may show stromal sclerosis and effacement of lung parenchyma. Pleural involvement or erosion of bronchial cartilage could additionally be seen; presence of these options favors lymphoma over a reactive course of. Histologic triad: (1) reactive lymphoid follicles with expanded marginal zones with/without follicular "colonization"; (2) polymorphous infiltrate with centrocyte-like and monocytoidlike cells; and (3) lymphoepithelial lesions. Polymorphous infiltrate: small lymphocytes, centrocyte-like cells, monocytoid-like cells, plasmacytoid lymphocytes, plasma cells, and few scattered large cells with vesicular chromatin and distinct nucleoli. Lymphoepithelial lesions: infiltration of 5 lymphoma cells into the respiratory epithelium.
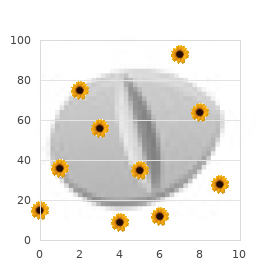
Generic ciriax 500 mg line
Uncoating is dependent upon actin and tyrosine kinases antibiotic bloating ciriax 250 mg cheap visa, possibly for motion of the capsid through the network of actin filaments antibiotic journal pdf 1000 mg ciriax order with amex, but not on dynamin, clathrin, caveolin, or flotillin (a marker protein for clathrin- and caveolin-independent endocytosis), endosome acidification, or microtubules. The tunnel opens on the base of the canyon and extends towards the 5-fold axis of symmetry. Because of the symmetry of the capsid, each virus particle might comprise as a lot as 60 lipid molecules. The lipids are thought to contribute to the stability of the native virus particle by locking the capsid in a stable conformation. The lipids are subsequently considered as switches, as a end result of their presence or absence determines whether or not the virus is stable or might be uncoated. The interaction of the virus particle with its receptor most likely initiates structural modifications in the virion that lead to the discharge of lipid. Uncoating in the Lysosome Most virus particles that enter cells by receptor-mediated endocytosis depart the pathway earlier than the vesicles attain the lysosomal compartment. However, these enzymes play an important function through the uncoating of members of the Reoviridae. The viral capsid is a double-shelled structure composed of eight completely different proteins. Infection of cells by reoviruses is sensitive to bafilomycin A1, an inhibitor of the endosomal proton pump, indicating that acidification is required for entry. Low pH activates lysosomal proteases, which then modify several capsid proteins, enabling the virus to cross the vesicle membrane. One viral outer capsid protein is cleaved and one other is removed from the particle, producing an infectious subviral particle. Attachment and Entry 147 A Virion Attachment Endocytosis Virion Endosome that interacts with membranes, on the surface. Consequently, subviral particles penetrate the lysosomal membrane and escape into the cytosol. Isolated infectious subviral particles cause cell membranes to become permeable to toxins and produce pores in synthetic membranes. These particles can provoke an an infection by penetrating the plasma membrane, getting into the cytoplasm instantly. Rather, viral particles and their components are transported by way of the actin and microtubule cytoskeletons. Such movement could be visualized in reside cells by utilizing fluorescently labeled viral proteins (Chapter 2). The cytoskeleton is a dynamic community of protein filaments that extends throughout the cytoplasm. It is composed of three forms of filament-microtubules, intermediate filaments, and microfilaments. Microtubules are organized in a polarized manner, with minus ends situated at the microtubuleorganizing center close to the nucleus, and plus ends positioned at the cell periphery. This association permits directed movement of cellular and viral parts over long distances. After the attachment of 1 protein to the cell receptor, the virus particle enters the cell by receptor-mediated endocytosis. The intact virion is composed of two concentric, icosahedrally organized protein capsids. The N terminus of 1N is modified with myristate, suggesting that the protein capabilities within the penetration of membranes. A pair of amphipathic -helices flank a C-terminal trypsin/chymotrypsin cleavage web site at which 1C is cleaved by lysosomal proteases. Dyneins and kinesins participate in movement of viral components during each entry and egress (Chapters 12 and 13). In some cases, the actin cytoskeleton is transformed throughout these processes, for example, when viruses bud from the plasma membrane. In the latter case, some form of the virus particle must bind on to the transport machinery. After leaving endosomes, the subviral particles derived from adenoviruses and parvoviruses are transported along microtubules to the nucleus. Although adenovirus particles have an general internet motion toward the nucleus, they exhibit bidirectional plus- and minus-end-directed microtubule movement. Adenovirus binding to cells prompts two different signal transduction pathways that enhance the online velocity of minus-end-directed motility. The signaling pathways are subsequently required for efficient supply of the viral genome to the nucleus. Adenovirus subviral particles are loaded onto microtubules via interplay of the capsid protein, hexon, with dynein. The particles transfer towards the centrosome and are then released and dock onto the nuclear pore complex, previous to viral genome entry into the nucleus. Some virus particles transfer alongside the surfaces of cells prior to entry till a clathrin-coated pit is encountered. If the cell receptor is rare or inaccessible, particles could first bind to extra ample or accessible receptors, similar to carbohydrates, after which migrate to receptors that permit entry into the cell. Filopodial bridges mediate cell-to-cell spread of a retrovirus in cells in tradition. The filopodia originate from uninfected cells and make contact with infected cells with their suggestions. The interplay of the viral envelope glycoprotein on the surface of infected cells with the receptor on uninfected cells stabilizes the interaction. Such transport is a consequence of actin-based motion of the viral receptor towards the uninfected cell. A number of different viruses enter the peripheral nervous system and spread to the central nervous system by way of axons. As no viral genome encodes the molecular motors or cytoskeletal buildings needed for long-distance axonal transport, viral adapter proteins are required to enable movement within nerves. After fusion on the plasma membrane, the viral nucleocapsid is carried by retrograde transport to the neuronal cell body. In distinction, different virus particles are carried to the nerve cell physique inside endocytic vesicles. The cytoplasmic area of the receptor engages the dynein mild chain Tctex-1 to permit retrograde transport of virus-containing vesicles. Virus-Induced Signaling by way of Cell Receptors Binding of virus particles to cell receptors not solely concentrates the particles on the cell surface, but also activates signaling pathways that facilitate virus entry and motion within the cell or produce cellular responses that improve virus propagation and/or have an effect on pathogenesis. Virus binding might lead to activation of protein kinases that trigger cascades of responses at the plasma membrane, cytoplasm, and nucleus (Chapter 14).
Order ciriax 250 mg overnight delivery
During poliovirus infection bacteria jekyll island 500 mg ciriax order amex, host cell translation is inhibited by 2 h after an infection and is replaced by translation of viral proteins antibiotic spacer ciriax 1000 mg otc. In this experiment, host translation was shut off by 5 h postinfection and was changed by the synthesis of viral proteins, some of that are labeled at the right. The first was found to be similar to a previously described protein, known as phosphorylated heat- and acid-stable protein regulated by insulin (Phas-I). This protein was known to be an necessary substrate for phosphorylation in cells treated with insulin and development elements. Phosphorylation of 4E-bp is carried out by a serine/threonine kinase, mammalian target of rapamycin kinase complicated 1 (mTorC1). Presence of development factors, oxygen, glucose, and energy lead to elevated translation as a outcome of phosphorylation of 4E-bp1 and ribosomal protein S6. As would be anticipated, mTorC1 is activated throughout infection, resulting in elevated protein synthesis under circumstances. Examples include inhibition of the tuberous sclerosis complicated (Tsc) by the human papillomavirus E6 protein and stimulation of phosphatidylinositol 3-kinase (Pi3k) by the adenovirus E4 Orf1 protein. Both herpes simplex virus kind 1 and human cytomegalovirus stimulate mTorC1 through inhibition of Tsc. The Us3 protein of herpes simplex virus type 1, a serine/ threonine kinase, functions as an Akt mimic and phosphorylates Tsc2. As anticipated, silencing of miR141 reduces the production of infectious virus particles. Binding of ligand to any one of several kinds of plasma membrane receptors initiates signaling to Pi3k related to the inner surface of the plasma membrane and phosphorylation and activation of this kinase. Shown are penalties of Akt activation that promote cell progress and proliferation by way of activation of the mTor kinase current in mTorC1. Activated mTor facilitates translation by a quantity of mechanisms and in addition induces autophagy. Instead, these 3 untranslated areas comprise a conserved sequence that binds the viral protein nsP3. These granules are believed to form when translation is inhibited by intracellular and extracellular stresses similar to nutrient deprivation or viral infection. Reduction in concentrations of both protein impairs formation of stress granules, and overproduction of either element stimulates formation of these aggregates. Late in viral infection, the viral proteinase 3Cpro cleaves G3bp1, disassembling stress granules, an occasion required for environment friendly viral replica. The presence of a noncleavable type of G3bp1 prevents the disassembly of stress granules and impairs viral copy. Stress granule elements may be redirected to different cellular sites in virus-infected cells. For example, the nsP3 protein of Semliki Forest virus sequesters G3bp1 into viral replication complexes. Removal of the nsP3 sequences which are necessary for interaction with this cellular protein impair viral manufacturing, suggesting a job for the protein in viral reproduction. Virus replication may lead to alteration of P bodies and redirection of their elements. The enterovirus 3Cpro proteinase cleaves a quantity of P-body parts including Xrn1, Dcp1a, and Pan3, disrupting P-body formation. In cells infected with the flavivirus West Nile virus, numerous P-body proteins are sequestered in viral replication complexes because the variety of P bodies diminish. Perspectives From the smallest to the largest, all viral genomes encode proteins that recruit the host cell translational equipment for manufacturing of proteins needed for viral copy. These viral proteins management or modify cellular translation proteins, ribosomes, and the signaling pathways that regulate their activities. Among all the viruses studied, every step of the interpretation process seems to be modified. The examine of such modifications has revealed a fantastic deal about how proteins are made and the way this process is regulated. Very early in an infection, intrinsic defense responses are mounted, and protein synthesis is inhibited in an try and restrict viral replica. Should infection proceed, mobile stress responses, which trigger further discount in translation, are activated. The interplay of cellular and viral modifications is a crucial determinant of the result of an infection. Studies of historical viral and cellular proteins that participate in translation reveal an evolutionary arms race as viral proteins change to overcome host defenses, and mobile proteins change in response. The outcomes reveal the remarkable plasticity of protein operate, and how genes and genomes have been shaped by challenges from viruses. When protein synthesis is inhibited as a response to stress, stalled translational complexes are routed to stress granules. Three marker proteins for stress granules, T cell intracellular antigen-1 (Tia-1), Tia-1-related protein (Tiar), and G3bp, are shown. Infection by some viruses, corresponding to West Nile virus, dengue virus, and poliovirus, might antagonize the formation of stress granules by interfering with the function of Tiar, Tia-1, or G3bp. G3bp may be cleaved by poliovirus 3Cpro, or sequestered into the replication complexes that type in cells contaminated with hepatitis C virus, Junin virus, alphaviruses, or flaviviruses, blocking stress granule meeting. The overwhelming majority of virus-induced modifications have an result on the initiation step of protein synthesis. Indeed, our detailed understanding of this step of translation has been a consequence of unraveling the consequences of viral infection. Although elongation and termination require far fewer mobile proteins, there are nonetheless examples of viral modulation of these steps as properly. An intriguing hypothesis is that the formation of stress granules is a part of an integrated response that includes intrinsic antiviral mechanisms. Emerging evidence indicates that intrinsic immunity and stress responses are linked at many ranges. How stress responses and intrinsic immunity work together at multiple levels might be a major goal of future analysis in this area. Poly(A) binding protein abundance regulates eukaryotic translation initiation factor 4F meeting in human cytomegalovirus-infected cells. Natural prevalence and characterization of two internal ribosome entry web site parts in a novel virus, canine picodicistrovirus, within the picornavirus-like superfamily. Because of the structural and useful compartmentalization of eukaryotic cells, parts of these particles are usually produced at a quantity of intracellular areas, and have to be introduced together for meeting. Intracellular trafficking and sorting of viral nucleic acids, proteins, and glycoproteins to the suitable websites is therefore an important prelude to the meeting of all animal viruses.
1000 mg ciriax buy with visa
Nongenomic Viral Nucleic Acid the presence of a viral nucleic acid genome has long been recognized as a definitive characteristic of virions antibiotics for bladder infection nitrofurantoin ciriax 1000 mg for sale. It is due to this fact tough to exclude the possibility that their presence is a functionally irrelevant and secondary consequence of nonspecific nucleic acid binding by viral structural proteins virus scanner free ciriax 250 mg cheap without a prescription. For instance, cellular glycoproteins will not be excluded from the membrane from which the viral envelope is derived. Furthermore, as a bud enlarges and pinches off throughout virus meeting, inside mobile elements may be trapped within it. As a result, preparations of those viruses may be contaminated with vesicles fashioned from mobile membranes. Indeed, analysis by the sensitive proteomic strategies offered by mass spectrometry has recognized from 50 to 100 mobile proteins in purified, enveloped particles of varied herpesviruses, filoviruses, and rhabdoviruses. Consequently, it might be troublesome to distinguish mobile elements particularly included into enveloped virus particles from those trapped randomly or copurifying with the virus. The mobile parts captured in retrovirus particles have been particularly well characterized. One of essentially the most uncommon properties of human immunodeficiency virus kind 1 is the presence of cellular cyclophilin A, a chaperone that assists or catalyzes protein folding. This protein is the most important cytoplasmic member of a ubiquitous family of peptidyl-prolyl isomerases. Cellular membrane proteins, corresponding to Icam-1 and Lfa1 (see Chapter 5), can additionally be incorporated in the viral envelope and can contribute to attachment and entry of retroviral particles. It is clear from these examples that virus particles contain a surprisingly broad repertoire of biologically active molecules which might be delivered to their host cells. This repertoire is undoubtedly larger than we presently respect, and the contributions of many parts of virus particles to the infectious cycles of many viruses have but to be established. This approach has additionally been invaluable in figuring out cellular proteins which might be also present, with considerable numbers detected in the particles of a number of families of enveloped viruses. The sizeable populations (50 to 100) of such mobile proteins emphasize the significance of distinguishing those proteins that contribute to viral reproduction from these included by chance. Purified extracellular herpes simplex virus kind 1 particles have been found by mass spectrometry to comprise forty nine cellular proteins. This set included proteins reported to be current within the particles of other herpesviruses, similar to cyclophilin A and actin, and many not detected previously. In these experiments, a phenotypically wild-type virus with a capsid protein fused to the green fluorescent protein was exploited to permit speedy and accurate measurement of yields of extracellular virus particles. Removal of 13 of these proteins from particles reduced virus yield considerably, even in cells that continued to synthesize the proteins. These observations established unequivocally that some mobile proteins integrated into herpesviral particles promote the subsequent cycle of replica. Perhaps viral reproduction is facilitated by supply of particular cellular proteins already associated with parts of viral particles or supply of the proteins to specific sites throughout entry. Comprehensive characterization of extracellular herpes simplex virus kind 1 virions. Two-step methodology to assess the importance of mobile proteins integrated into herpes simplex virus type 1 particles. Now that many constructions of particles or their elements have been examined, we can appreciate the surprisingly diverse architectures they exhibit. Nevertheless, the simple principles of their construction proposed greater than 50 years in the past stay pertinent: with few exceptions, the capsid shells that encase and protect nucleic acid genomes are built from a small variety of proteins arranged with helical or icosahedral symmetry. The detailed views of nonenveloped virus particles provided by X-ray crystallography emphasize simply how properly these protein shells provide safety of the genome during passage from one host cell or organism to one other. They have also identified a number of mechanisms by which equivalent or nonidentical subunits can work together to type icosahedrally symmetric structures. More-elaborate virus particles, which can contain extra protein layers, a lipid envelope carrying viral proteins, and enzymes or different proteins necessary to initiate the infectious cycle, pose larger challenges to the structural biologist. Indeed, for a couple of years we possessed only schematic views of these buildings, deduced from negative-contrast electron microscopy and biochemical or genetic strategies of analysis. In the intervening period of simply 5 years, these strategies have attained atomic-level resolution, offering exceptional views of enormous viruses with a number of components, viral envelopes, and, in some instances, the group of genomes within particles. The structural descriptions of ever-increasing numbers of viruses representing various families have additionally allowed unique insights into evolutionary relationships among seemingly disparate viruses or viral proteins. The more just lately described giant viruses, similar to pandoravirus, with particles so large that they can be seen by gentle microscopy, additionally pose new technical challenges and counsel that unanticipated structural principles remain to be elucidated. Reconstructing virus structures from nanometer to near-atomic resolutions with cryo-electron microscopy and tomography. The constructions of human rhinovirus and Mengo virus: relevance to operate and drug design. Structure of a common chilly virus and useful relationship to other picornaviruses. Three-dimensional construction of the human immunodeficiency virus type 1 matrix protein. Insight into the mechanism of the influenza A proton channel from a construction in a lipid bilayer. Structure of the haemagglutinin membrane glycoprotein of influenza virus at 3 � resolution. The fusion glycoprotein shell of Semliki Forest virus: an icosahedral meeting primed for fusogenic activation at endosomal pH. Cryo-electron microscopy reveals the useful group of an enveloped virus, Semliki Forest virus. Proteomic and biochemical analysis of purified human immunodeficiency virus sort 1 produced from contaminated monocyte-derived macrophages. The bodily properties of the virion are obstacles to this seemingly easy aim. Furthermore, the viral genome is encapsidated in a steady coat that shields the nucleic acid as it travels by way of the tough extracellular setting. These impediments should all be overcome through the process of viral entry into cells. Encounter of a virus particle with the floor of a prone host cell induces a collection of events that result in entry of the viral genome into the cytoplasm or nucleus. The first step in entry is adherence of virus particles to the plasma membrane, an interplay mediated by binding to a selected receptor molecule on the cell surface. The receptor performs an important position in uncoating, the process by which the viral genome is uncovered, so that gene expression and genome replication can start. Interaction of the virus particle with its receptor might provoke conformational adjustments that prime the capsid for uncoating. Alternatively, the receptor may direct the virus particle into endocytic pathways, the place uncoating could also be triggered by low pH or by the action of proteases. The genomes of viruses that replicate within the nucleus are moved to that location by cellular transport pathways.